以前は異なる学問として考えられていた免疫学分野と代謝学分野において、代謝プロセスが免疫細胞の機能を巧妙に制御しているという新たな認識が広まりつつあります。免疫代謝学として知られるこの分野は、代謝と免疫機能の複雑な相互作用を探求し、病原体や疾患、治療法への身体の応答の元となる、込み入った網のような仕組みを解明するのに役立ちます。現在の研究では、代謝プロセスが免疫細胞の発達や活性化、機能などにどのような影響を与えるかの理解を試みており、最終的には新たな治療標的の特定を目指しています。
がん微小環境 (TME) における代謝と免疫機能の相互作用は、腫瘍の進行や免疫系による監視の回避、免疫療法への応答に直接的に影響するため、特に重要です。本ブログでは、解糖系とTCAサイクル間の代謝チェックポイントがどのように免疫応答と炎症を制御しているかの理解に役立つ、腫瘍細胞や免疫細胞におけるグルコース代謝に関して現在までに分かっている知見を概説します。
また、ブログの最後には、TMEにおける免疫代謝の研究に関連するCST製品リストを掲載しています。
<Jump to the immunometabolism target product list>
がん微小環境における免疫代謝
がんの塊は、がん細胞と、多様な非がん細胞(様々なタイプの免疫細胞を含む)で構成されています。血管やシグナル伝達分子、細胞外マトリクスなども含むこれらの不均一な細胞集団は、TMEとして知られる複雑な生物システムを構築します。
免疫細胞は、TME内でがんの進行の調節に関わる重要な役割を担います。「免疫系による破壊の回避」と「腫瘍による炎症の促進」は、がんの特性のうちの2つですが、これらはTMEにおける免疫細胞の役割を浮き彫りにしています。同時に、免疫細胞の代謝は、免疫細胞の分化や表現型の変換、エフェクター機能に重要であるため、免疫細胞内における代謝の変化は免疫代謝学分野において不可欠な要素です。
さらに、組織の脈管形成や、栄養および酸素を利用できるかどうか、免疫細胞と他の細胞タイプとのクロストークなど、細胞外の因子がTMEにおける免疫細胞の代謝活性にさらなる影響を与えます。特に、組織内の免疫細胞の配置は、その代謝活動に重大な影響を与えます。
グルコース代謝と代謝チェックポイント
グルコースは、解糖系の重要な構成因子であり、細胞内の10ステップの代謝経路を用いてグルコースをエネルギーに変換します。グルコースは、Glucose transporter 1 (Glut1、SLC2A1) により細胞内に取り込まれた後、細胞質で解糖系により分解され、ピルビン酸やATP、NADHを生成します。続いて、酸素が十分にある細胞では、ピルビン酸はミトコンドリアのマトリックスに移動し、そこでPDC (Pyruvate dehydrogenase complex) によりアセチルCoAに変換されます。アセチルCoAはその後、TCA (Tricarboxylic acid) サイクルに入り、そこでNADHとFADH2を生成します。NADHとFADH2は、ミトコンドリア内膜中の電子伝達鎖中で酸化され、ATPを生成します。
関連ブログ: 解糖系とは何か?代謝におけるその役割
一方、酸素が不足している細胞では、PDH (Pyruvate dehydrogenase) 活性が阻害されることにより、解糖系で生成されたピルビン酸はLDHA (Lactate dehydrogenase) を介して乳酸産生に回されます。PDH活性は、PDHK (Pyruvate dehydrogenase kinase) によるリン酸化を介して調節されています。また、PDH活性は、解糖系やTCAサイクルの相対的な活性に影響を与えます。そのため、PDHは、酸素レベルの違いに応じて細胞のエネルギー産生と代謝に影響を与える、制御された代謝チェックポイントタンパク質であると考えられます。
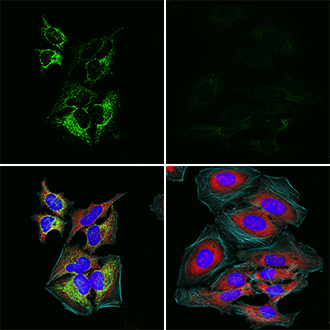
モック処理したHeLa細胞(左)または5 mM Sodium dichloroacetate (PDHK) 阻害剤で16時間処理しSer293のリン酸化を抑制したHeLa細胞(右)を、Phospho-Pyruvate Dehydrogenase α1 (Ser293) (E4V9L) Rabbit mAb #37115(緑)とPhospho-S6 Ribosomal Protein (Ser235/236) (E2R1O) Mouse mAb #62016(赤)を用いて免疫蛍光染色し解析しました。
がん細胞において、酸素が豊富な環境下でピルビン酸が乳酸産生に回されることを、好気的解糖またはワールブルク効果と呼びます。この現象は、増殖性の代謝の特性であり、がん細胞が代謝をリプログラミングしていることを示しています。これにより、がん細胞は、十分に酸素がある状態であってもエネルギー産生に解糖系を活用できます。そのため、多くのタイプのがん細胞における増殖と生存率が向上すると考えられています。
リソース: ワールブルク効果経路の概略図
免疫とがんにおける代謝チェックポイントの役割
先述したとおり、解糖系とTCAサイクル間の代謝チェックポイントは、細胞機能、特に適応免疫応答やがんの進行の制御に極めて重要な役割を果たしています。
以下では、この代謝チェックポイントが、様々なタイプの免疫細胞とその機能、またはがん細胞の代謝にどのような影響を与えるかを解説します。
- 自然免疫におけるM1様マクロファージとM2様マクロファージの活性化の制御の違い:M1様マクロファージの極性化には、PDHK (Pyruvate dehydrogenase kinase) によるPDH (Pyruvate dehydrogenase) のリン酸化が必要です。極性化プロセスでは、PDHK1やPDHK3、PDHK4の発現が上昇し、PDHのリン酸化やその活性の阻害につながります。この代謝の変化は、ミトコンドリアでの酸化的リン酸化から細胞質での解糖へと細胞を転換させます。PDHK1のノックダウン、あるいはPDHK2とPDHK4の両方の欠失は、解糖系の活性を低下させてM1への極性化を抑制する一方で、ミトコンドリアにおける呼吸を増加させてM2への極性化を増加させます1-3。
- 適応免疫よるCD4陽性エフェクターT細胞 (Teff) と制御性T細胞 (Treg) の増殖や分化、生存率の様々な制御:活性化したCD4陽性T細胞は、エフェクターT細胞 (Teff) と制御性T細胞 (Treg) に分化します。モデルマウスにおけるCD4陽性T細胞集団の研究から、PDHK1は、制御性T細胞よりもエフェクターTh17細胞において高発現していることが分かっています。PDHK1活性の阻害またはPDHK1発現の下方制御は、解糖系を抑制し、エフェクターTh17細胞の増殖や分化、生存率を低下させます。一方、PDHK1活性の阻害ままたはPDHK1の発現を下方制御すると、ミトコンドリアにおけるTCAサイクルと酸化的リン酸化が促進され、制御性T細胞の分化が促進されます。これらの結果から、エフェクターTh17細胞と制御性T細胞は、増殖や分化、生存率のために、それぞれが解糖系とミトコンドリアにおける呼吸を異なる形で必要としていることが分かります4。
M1様マクロファージやエフェクターTh17細胞などの炎症性免疫細胞は、より多くのグルコースを取り込み、好気的解糖を亢進します。対照的に、M2様マクロファージや制御性T細胞などの抗炎症性免疫細胞は、酸化的リン酸化をより多く行います5。
- がん細胞における好気的解糖:がん細胞は、酸素が十分にある場合でも大量のグルコースを取り込み、解糖系を介してピルビン酸に変換し、続いて乳酸発酵を行います (好気的解糖、ワールブルグ効果)。好気的解糖は、生合成中間体やエネルギー (ATP) を増やし、NADPHなどを減らして、がん細胞増殖のための高分子の生合成をサポートします。がん細胞では、PDHK1の発現と活性は制御されています。例えば、PDHK1の発現は、HIF-1αとc-Mycにより上方制御されます。さらに、いくつかのがん細胞においてPDHK1は、様々な発がん性チロシンキナーゼによりTyr243がリン酸化され、酵素活性の上昇につながることが分かっています。増大したPDHK1活性は、PDHK1活性の亢進は、解糖系への代謝転換を促進します6。
以下の図は、ヒトおよびマウスの免疫細胞における免疫代謝の研究に使用できる、マクロファージおよびT細胞マーカーを示しています。
 完全なHuman Immune Cell Marker Guide(ヒト免疫細胞マーカーガイド)をご覧になって、ヒト免疫系中の追加的な細胞タイプを探索してください。
完全なHuman Immune Cell Marker Guide(ヒト免疫細胞マーカーガイド)をご覧になって、ヒト免疫系中の追加的な細胞タイプを探索してください。

マウスの免疫系におけるその他の細胞タイプについては、免疫細胞マーカーガイド(マウス) の完全版をご覧ください。
免疫代謝が重要な理由
細胞の代謝は、免疫細胞の機能を制御する上で重要な役割を担っています。免疫代謝という新たな分野の研究は、代謝が免疫系にどのような影響を与えるかについて、新たな視点を提供することが期待されています。
免疫代謝研究用のCST抗体製品
T細胞マーカー |
|
| Th17 | Treg |
| IL-17A (D1X7L) Rabbit mAb #13838 | FoxP3 (D6O8C) Rabbit mAb #12632 |
| IL-17RA (D1Y4C) Rabbit mAb #12661 | IL-2Rα/CD25 (D6K5F) Rabbit mAb #13517 |
| IL-17F (D3M4D) Rabbit mAb #13186 | |
参考文献:
- Jeon JH, Thoudam T, Choi EJ, Kim MJ, Harris RA, Lee IK. Loss of metabolic flexibility as a result of overexpression of pyruvate dehydrogenase kinases in muscle, liver and the immune system: Therapeutic targets in metabolic diseases. J Diabetes Investig. 2021;12(1):21-31. doi:10.1111/jdi.13345
- Tan Z, Xie N, Cui H, et al. Pyruvate dehydrogenase kinase 1 participates in macrophage polarization via regulating glucose metabolism. J Immunol. 2015;194(12):6082-6089. doi:10.4049/jimmunol.1402469
- Min BK, Park S, Kang HJ, et al. Pyruvate Dehydrogenase Kinase Is a Metabolic Checkpoint for Polarization of Macrophages to the M1 Phenotype. Front Immunol. 2019;10:944. Published 2019 May 7. doi:10.3389/fimmu.2019.00944
- Gerriets VA, Kishton RJ, Nichols AG, et al. Metabolic programming and PDHK1 control CD4+ T cell subsets and inflammation. J Clin Invest. 2015;125(1):194-207. doi:10.1172/JCI76012
- Pålsson-McDermott EM, O'Neill LAJ. Targeting immunometabolism as an anti-inflammatory strategy. Cell Res. 2020;30(4):300-314. doi:10.1038/s41422-020-0291-z
- Hitosugi T, Fan J, Chung TW, et al. Tyrosine phosphorylation of mitochondrial pyruvate dehydrogenase kinase 1 is important for cancer metabolism. Mol Cell. 2011;44(6):864-877. doi:10.1016/j.molcel.2011.10.015
サイエンティフィックマーケティングライターであり、CSTのブログマネージャーであるAlexandra Foleyが、本ブログ記事の執筆に協力しました。24-HMC-64201
24-HMC-64201