上皮間葉転換 (EMT) は、分化した上皮細胞が間葉系細胞に近い性質をもった細胞に形質を変えるプロセスであり、1980年代にハーバード大学の先駆的な生物学者であるElizabeth “Betty” Hayによって発見されました。
革新的な電子顕微鏡技術を用い、Hayはニワトリ胚の原子線条の上皮細胞が「逆の」発生プロセスを経るという驚くべき発見をしました。これは、分化は一方向のみに進むという、広く受け入れられていた概念に反するものでした。これらの「脱分化」した上皮細胞は、紡錘状の運動性の高い細胞に形質を変化させ、起源の部位から移動して遠位の組織の形態形成に寄与することが分かりました。Hayは表現型の可塑性を端的に示すこの現象を「上皮間葉転換 (EMT:epithelial-to-mesenchymal transformation)」と名付け、その後の綿密な解析により、これは動物の発生において重要かつほぼ普遍的な現象であることが明らかになりました[1]。
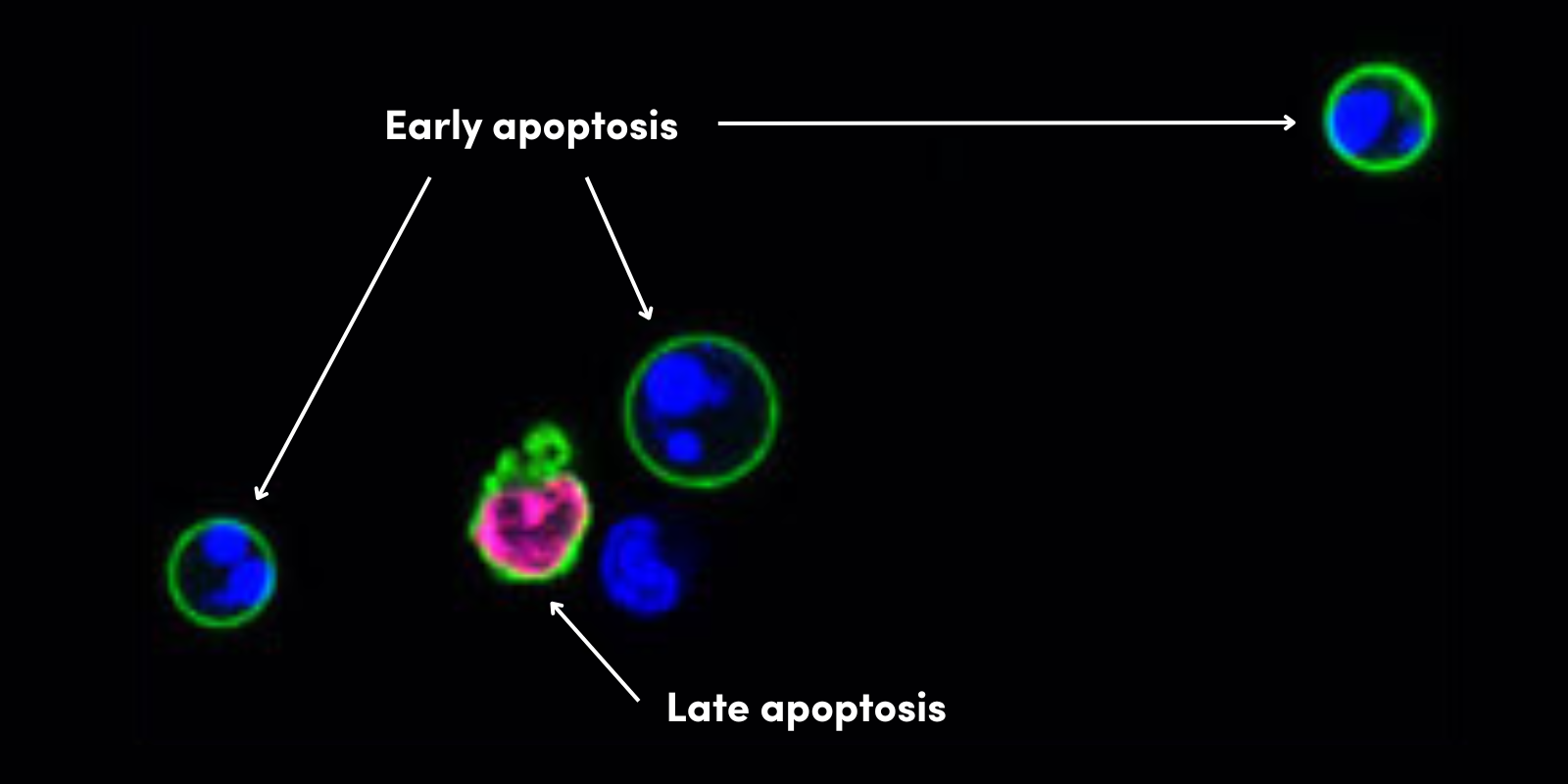
EMTのプロセスにおいて細胞は一連の表現型の変化を示し、これらの多くはEMTの特性として認識されています (PMID:19487818)。これらの変化の代表例として、細胞極性の消失、アクチン細胞骨格の脱分極、近隣の細胞に結合する結合複合体の解体などが挙げられます。これによって間葉様の形質を得た細胞は特徴的な形態をとり、主に接着斑を介して近接細胞や基質と相互作用し、さらに細胞外マトリクスの成分と相互作用することで運動性が促進されます。
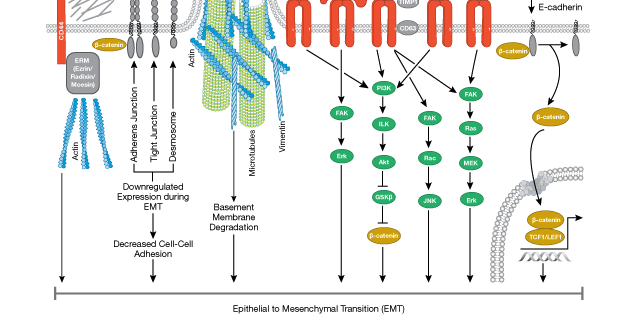
その後の生化学的研究や遺伝学的研究によって、特定の遺伝子の発現変化がこれらの表現型の変化に関与していることが明らかにされています。例えば、一連の転写抑制因子(SNAIL、TWIST、ZEB1など) の発現が増加することで、上皮の表現型を維持するために必要な細胞接着タンパク質 (E-Cadherin、Desminなど) の発現が低下します。同様に、間葉の性質に関与する遺伝子 (α-SMA、Vimentin、FSP1/S100A4など) の選択的な発現増加や、細胞の遊走を促進する細胞外マトリクス関連因子 (Fibronectin、Vitronectinなど) の発現増加がみられます。上記のEMT転写因子の発現は細胞の内外のシグナル伝達パスウェイによって制御されており、これらのパスウェイの多くは正常な動物の発生において重要な役割をもつことが知られています。細胞外のパスウェイにはTGF-β、Wnt、FGFおよびHedgehogパスウェイ があり、これらは古典的な細胞内シグナル伝達経路 (ERK、 MAPK、PI3K/AKT、Ras、SMADsなど) を介して作用します。
また、EMTは生理的な意義の異なる3つのタイプに分類することができます。タイプⅠは上記のように正常な胚発生の一部として起こる現象、タイプⅡは創傷治癒や線維症のプロセスに関連して起こるもの、タイプⅢは腫瘍細胞の転移能の獲得に寄与すると考えられる脱分化プログラムです (PMID:18485877)。胚発生期における細胞の表現型の可塑性は比較的受け入れ易いですが、分化の決定づけがなされた後の上皮細胞の脱分化や、タイプⅡやタイプⅢのような疾患の進行に寄与するEMTについては、現在も盛んに議論が続けられています (PMID:26784242)。特に後者については、Hayによって最も「恐ろしい」EMTの側面として紹介されており (PMID:8714286)、EMTの発見以来、恐らく最も盛んに議論されてきました (PMID:27042694)。
さて、この細胞のアイデンティティ・クライシスは実際にがんの進行に関与しているのでしょうか?生体内でEMTを観察するのは困難で、この疑問に結論を下すのは容易ではありません。 また、通常転移巣は原発巣の表現型が再現されることが分かっており、間葉から上皮への再分化が必要であると考えられてきました。初期の腫瘍形成の原因となるジェネティック、エピジェネティックな異常を考慮すると、このような一時的かつ可逆的でありながら部分的に制御された可塑性はありそうにないと考える人もいます (PMID:23201165)。また、意図的にEMTを誘導してもがん転移の誘導はみられなかったという知見や、標準的なEMT遺伝子は発現することなく転移がみられたという研究成果もあり、EMTはがん転移の必須条件とは言えないようです (PMID:26560033)。
しかし、近年のWeinberg博士のグループの研究成果から、いくつかの研究でEMTと転移の関係性が見出されなかった理由を説明できる可能性があります。近年のWeinberg博士のグループの研究成果から、いくつかの研究でEMTと転移の関係性が見出されなかった理由を説明できる可能性があります。その中の研究で、乳がん細胞の腫瘍形成能は上皮と間葉の中間である「ハイブリッド」状態で最大化し、完全に間葉を誘導すると腫瘍始原細胞としての機能が低下することが示されました (Krögeret al. 2019、PMID:30910979)。Weinberg博士はさらに2019年のアメリカがん学会 (AACR) 年会で、この「ハイブリッド (E/M)」状態の細胞はより効率的に免疫系による検出を回避し、腫瘍形成能をさらに高めている可能性があるという、非常に興味深い知見を報告しました。
がんや線維症といった疾患の進行とEMTの関係については解明すべき課題が多く残されています。Betty Hayの革新的な発見は、将来的に細胞の表現型の可塑性に基づいたより効果的な治療法の開発に繋がることが期待されています。
EMTのシグナル伝達の詳細は、CSTのパスウェイ図をご覧ください。
[1] EMTは可逆的な変化であり、後に間葉上皮転換 (MET) というプロセスを受けることが明らかにされました。これを踏まえ、epithelial-to-mesenchymal transformationという用語は後に、epithelial-to-mesenchymal transitionと改められました。これは悪性形質転換 (malignant transformation) との混同を防ぐためと、EMTのもつ可塑性をよりよく表すためです。