細胞老化とは、細胞が代謝的な活性を保持したまま細胞周期から不可逆的に離脱し、増殖刺激への応答性を失った状態を指します。老化した細胞は、がん、糖尿病、加齢といった多くの生理的、病理的プロセスに影響を及ぼします。したがって、細胞老化がこれらの現象にどのように関わっているかを明らかにすることで、細胞老化の促進や抑制を介した新たな疾患の治療法が開発できる可能性があります。
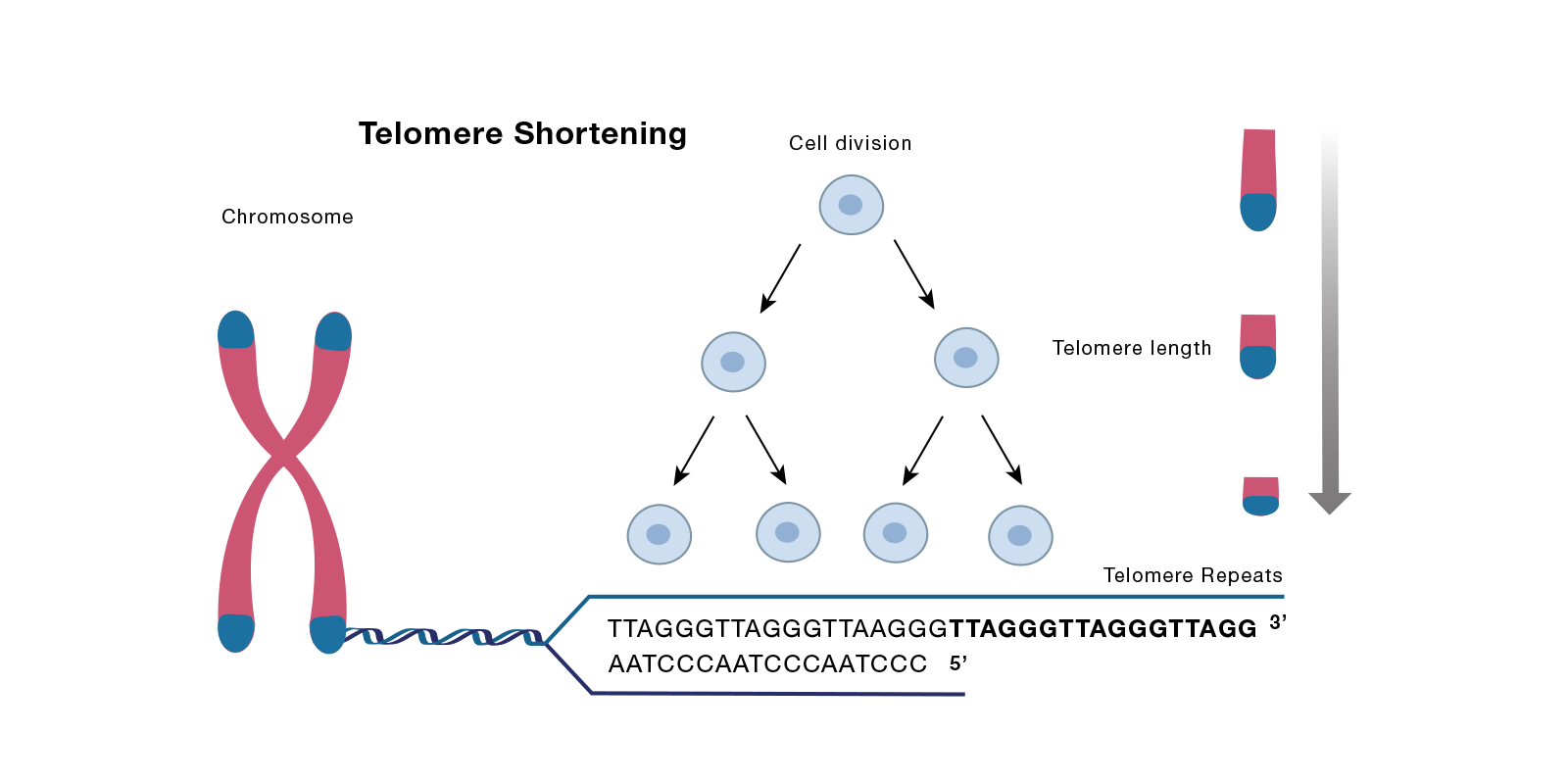
様々な細胞外シグナルによるDNA損傷や、細胞内のがん遺伝子の活性化が、細胞老化の主要な誘導因子となります。一方、正常なテロメアDNAの短縮も細胞老化の原因となります。数十年前に、Leonard Hayflick博士とPaul Moorehead博士が、in vitroで増殖した細胞は、一定回数の細胞分裂を経た後に細胞周期を停止させる現象を報告し、現在この現象は複製老化と呼ばれています (Hayflick and Moorhead, 1961; PMID: 13905658). その後の研究で、この細胞分裂の限界は、テロメアと呼ばれる染色体の末端構造の短縮に由来するシグナルによって部分的に制御されていることが明らかになりました。テロメアの短縮は、細胞分裂に先行する正常なDNA複製の各サイクルで起こります [(Bodnar AG, 1998; PMID 9454332) または次の総説(Kuilman T 2010, PMID 21078816)]。
テロメアは、数百から数千の短いヌクレオチド反復配列で構成されており、最末端ではGTに富む単鎖のオーバーハングを形成しています。幾つかのテロメア結合タンパク質がこれらの配列を認識して結合し、Shelterin複合体と呼ばれる複合体を形成します。この複合体は染色体末端を分解から保護するキャップ構造の形成を促進します。Shelterin複合体は、TRF2 (Telomere repeat factor 2)、TERF2IP (Telomeric repeat-binding factor 2-interacting protein、RAP1とも呼ばれます) 、TPP1 (Telomere protection protein) を含む複数のタンパク質で構成されます。末端複製問題 (染色体は直鎖DNAであり、複製の際に末端を完全にコピーできないという現象) のため、細胞分裂が起こるたびにテロメア配列の長さは短縮されます。これらの配列が臨界長に達すると、細胞のDNA修復機構が活性化して細胞老化が引き起こされ、細胞分裂が停止します。細胞は、テロメア配列を伸長するテロメラーゼと呼ばれる酵素を発現してテロメアの短縮を防ぎます。しかし、加齢により、あるいはテロメラーゼ活性の低いタイプの細胞では、複製老化が起こります。
がん細胞の多くは複製老化を逃れ、無制限に増殖しますが、テロメラーゼの発現や活性の亢進がこの一因になっています。テロメラーゼは、DKC1 (dyskerin) を含むタンパク質複合体と協調して作用し、細胞分裂の際に新規にテロメア末端配列を合成します。 MRN (Mre11-Rad50-Nbs1) 複合体も、テロメア安定性の維持に重要な役割を果たし、この複合体をコードする遺伝子の変異が、幾つかのがんのリスク増大に関与しています。
主にテロメア短縮によって引き起こされる複製老化により、加齢とともに織中に老化細胞が蓄積します。したがって、老化細胞を選択的に標的とする薬剤や、テロメラーゼ活性を高めて複製老化を制限する薬剤が、加齢による弊害を抑制してヒトの寿命を延長し、治療的な価値を持つ可能性があります。実際にテロメアは細胞時計として機能することから、その分解を遅延させることで細胞老化を制限し、組織の健康と機能を延長できる可能性があります。
複製老化の起こった細胞は、DNA損傷などによって誘導される細胞老化と同様の多くの形態的、代謝的な特徴を示します。したがって、細胞周期停止を調節するタンパク質の発現変化や、SASP (senescence-associated secretory phenotype) と呼ばれる表現型の獲得を検出するアッセイにより、様々な実験モデルにおいて老化細胞の存在を検出することができます。
テロメアと細胞老化の関係について、詳しくはCST 細胞老化シグナル伝達経路をご覧ください。