CSTは、PD研究を前進させるためにMichael J. Foxパーキンソン病財団 (MJFF) とパートナーシップを締結したことを大変嬉しく思います。本パートナーシップの詳細をご覧になり、PDのリソースを探索してください。
—
ヒトの細胞は、内外のストレスにさらされても細胞の健康が保たれるように、非常に複雑な品質管理システムを持っています。このシステムには、デブリ (細胞片) や折り畳みに異常があるタンパク質を除去し、有害な細胞成分を管理する、複雑で高度に制御された複数の経路が含まれます。パーキンソン病 (PD) などの多くの一般的な神経疾患では、加齢とともに損なわれることもあるこれらの複雑な品質管理を行う経路が、様々なメカニズムにより障害を受けていることが同定および明らかにされています。
特にPDは、ドーパミン作動性神経細胞の機能不全による、協調運動を司る神経伝達物質であるドーパミンの減少を特徴とします。孤発性および遺伝性PDのいずれにおいても、この機能不全は、オートファジーやリソソームによる分解に関与する様々なタンパク質の障害が原因の1つであり、その結果、折り畳みに異常があるタンパク質が蓄積し、最終的にドーパミン作動性神経細胞が死に至ると考えられています。こうした神経細胞の喪失は、PD患者に特徴的な振戦の原因となります。
インタラクティブパスウェイ図:オートファジーシグナル伝達
近年では、オートファジーやマイトファジー、エンドリソソームの輸送などの細胞の健全性や恒常性を維持するための主要なプロセスの機能障害や機能低下が、治療介入の焦点となりつつあります。遺伝性PDおよび孤発性PDの発症機序をより深く理解することは、新たな治療法への道を切り開くことにつながります。
本ブログ記事では、神経細胞の恒常性を維持するために必要なオートファジーやマイトファジー、エンドリソソームシステムの概要を紹介します。また、これらの経路における障害がどのようにパーキンソン病に寄与するかを解説し、さらに、近年発表された科学論文を参照しながら、パーキンソン病におけるリソソーム障害の研究に使用されている最新の手法を紹介します。
<<リソソームの機能不全の研究用の手法のセクションに飛ぶ >>
神経細胞の恒常性維持におけるオートファジー、マイトファジーおよびエンドリソソームシステム
PDの発症機序と進行を理解するためには、健康な神経細胞におけるオートファジー、マイトファジー、エンドリソソーム経路の役割を認識することが不可欠です。神経細胞は、有糸分裂を終えた分泌細胞であるため、その恒常性を維持することは特に重要であり、神経細胞では軸索と細胞体の間の距離が遠いことことから、これらの経路の重要性がさらに高まります。
.png?width=780&height=338&name=23-EMG-00600-Lysosomal%20%26%20Mitochondrial%20Dysfuntion%20in%20PD%20Blog_Fig%201%20(1).png) エンドリソソーム経路とオートリソソーム経路が正しく機能している場合、折り畳みに異常があるタンパク質や損傷したオルガネラを細胞から除去します。
エンドリソソーム経路とオートリソソーム経路が正しく機能している場合、折り畳みに異常があるタンパク質や損傷したオルガネラを細胞から除去します。
オートファジーとエンドリソソームの輸送は、折り畳みに異常がある、または凝集しやすいタンパク質や、機能不全のミトコンドリアなどの損傷したオルガネラを効率的に除去することにより、細胞の恒常性の維持を可能にします。
細胞の基本:オートファジー
細胞の主な分解経路の1つであるオートファジーは、細胞質成分を二重膜のオートファゴソーム内に選択的または非選択的に取り込むことで、損傷したオルガネラと折り畳みに異常があるタンパク質を除去する、進化的に保存された異化プロセスです。神経細胞におけるオートファゴソームの生合成は、末梢の軸索とシナプスにおいてのみ開始され、末梢で常に必要とされる新しいタンパク質とオルガネラの補充に役立ちます1。
神経細胞におけるオートファジー:神経伝達物質の再利用とオートファジーのバランスの維持は、活性依存的な制御機構です。発達中の神経細胞では、オートファジーは樹状突起スパインの刈込みに必要であり、一方成熟した神経細胞、特にシナプス小胞では、細胞質およびシナプス前タンパク質の選別と代謝回転に重要なプロセスです。
細胞の基本:マイトファジー
マイトファジーは、損傷した、または余分なミトコンドリアをオートファジーによって選択的に分解することを指し、健全なミトコンドリアネットワークを維持するために重要です。神経細胞では、マイトファジーは細胞体と軸索遠位部の両方で生じます。
神経細胞におけるマイトファジー: ストレス状態にある神経細胞内では、PINK1タンパク質とE3ユビキチンリガーゼであるParkinが、損傷したミトコンドリアを標的としてユビキチン化します。その結果、オートファジーを誘導し、p62/SQSTM1やOptineurinなどのユビキチン結合オートファジー受容体がリクルートされます。しかし、PINK1またはParkinの欠損は、 in vivoでは基本的なマイトファジーに影響を与えないことが示されており2,3、機能不全のミトコンドリアを除去する、PINK1/Parkinから独立した機構が存在する可能性があります。
細胞の基本:エンドリソソーム経路
すべての細胞タイプに存在するエンドリソソーム経路は、細胞内に持ち込まれた様々な膜カーゴの選別、調節、リサイクルを行う一連の動的なオルガネラです。このプロセスでは、AP2で覆われたエンドサイトーシス小胞が、形質膜からカーゴを取り込み、リサイクルエンドソーム (マーカー:Rab35、Rab11) を介して形質膜に戻すか、分解経路に移行させます。小胞を覆っていたAP2が除去されると、リソソーム分解の標的となる分子を含むエンドサイトーシス小胞は、初期エンドソーム (マーカー:Rab5、EEA1) と融合し、後期エンドソーム (マーカー:Rab7) へと成熟します。神経細胞のエンドリソソーム経路では、後期エンドソームはシナプス終末でオートファゴソーム (マーカー:ATG8、LC3) と融合します。エンドソームは、ダイニンモーターを用いて微小管に沿って徐々に移動し、酸性化した後、細胞体のリソソームと融合してカーゴを分解します。
神経細胞内のエンドリソソーム輸送:後シナプスでは、エンドリソソーム経路はシナプス活性に依存して、膜貫通型受容体であるAMPARとGABAaのリサイクルや分解に関与しています。このプロセスにおいて、リソソームは活性依存的に輸送され、シナプスの活性化によって樹状突起スパインにリクルートされ、そこでAMPARとGABAaを分解します。この動的なプロセスは、神経可塑性と神経伝達物質の放出を可能にし、維持するために不可欠です。
パーキンソン病の標的:PARK遺伝子
PDの多くは孤発性であり、遺伝的要素を持つものは10%未満です。しかし、遺伝形式をもつPARK遺伝子は、PDの生物学を解明する上で極めて重要です。PDのリスク遺伝子の多くは、恒常性の維持に重要であり、エンドリソソーム経路において役割を担うα-Synuclein、LRRK2、VPS35、Parkin、PINK1、およびDJ1のタンパク質をコードする遺伝子があります4。
CSTは、オートファジーからエンドリソソーム経路まで、PDのリスク遺伝子から産生される機能性タンパク質の研究用の抗体を提供しています。
- PDのリスク遺伝子である、オートファジーとエンドリソソーム経路に関与する遺伝子:
- リソソーム酵素:Cathepsin B
- リソソーム酵素: Glucocerebrosidase (GCase/GBA)
- 液胞型ATPaseサブユニット:ATP6V0A1
- リソソームのカリウムチャネル: TMEM175
- 遺伝性PDと関連する、オートファジーとエンドリソソーム経路に含まれる遺伝子:
- Synaptojanin-1
- LRRK2
- リソソームのATPase: ATP13A2 (PARK9)
- PINK1
- Parkin
オートファジーとリソソームの障害がどのようにPDに寄与するか:α-synの例
α-Synuclein (α-syn) は、脳内で最も多く存在するタンパク質の1つであり、その発現は可塑性に関与していることを示しています5。 健康な神経細胞で正しく機能している場合、α-synはシナプス前タンパク質として、神経伝達物質の放出に役割を果たしていると考えられています。このタンパク質は、翻訳後に広く修飾される、もともとは構造化されていないタンパク質であるため、存在する細胞環境に応じて多くの異なる立体配座をとると考えられています6。
折り畳みに異常があるα-synの、レビー小体と呼ばれるタンパク質封入体への蓄積と凝集は、パーキンソン病の発症と進行における1つのステップであると考えられています。疾患状態では、α-synは可溶性のオリゴマー中間体を形成し、レビー小体に見られる不溶性のアミロイド線維に成熟することで凝集体を形成します。オリゴマーのα-syn (o-α-syn) は、有害な物質であると考えられています。α-synの凝集体は、一度形成されると、細胞間の伝播を通じて、プリオン様様式で中枢神経系全体に広がる可能性があります7,8,9。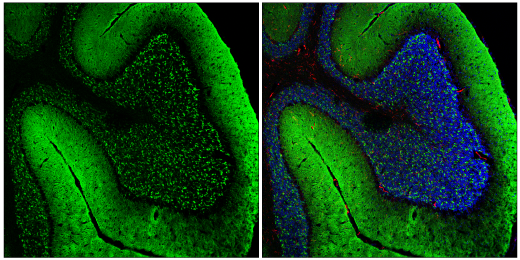
固定済みのマウス凍結小脳組織を、α-Synuclein (E4U2F) XP® Rabbit mAb #51510 (左、緑) で免疫蛍光染色し、F4/80 (BM8.1) Rat mAb #71299 (右、赤) およびDAPI #4083 (右、青) で共標識し、共焦点顕微鏡で解析しました。
通常、α-synはオートファジーによって除去されますが、その凝集体は、ATG7を抑制してmTORレベルを上昇させることにより、オートファジー-リソソーム経路を直接変化させ、オートファジーの開始を阻害する可能性があります。凝集したα-synは、遺伝性PDと孤発性PDの両方において、自身の分解という必要性が高い要求に応えるため、オートファゴソームの形成を促進すると考えられていますが10、その過剰発現はオートファゴソームの生合成を阻害することも示されています11。 ドーパミンで修飾されたα-synが、このプロセスをさらに増悪させることが示されており、これがPDにおけるドーパミン作動性神経細胞の選択的脆弱性の理由である可能性があります12。
パーキンソン病においてα-synucleinが分解を回避する方法
- ある研究では、α-synのA53TおよびA30P変異体は、in vitroにおいて、シャペロン介在性オートファジー経路による自身や他の基質の分解のための、リソソームによる取り込みを阻止しました13。
- また、α-synの凝集体は、オートファジー小胞の逆行性輸送を阻害し、オートファゴソームの成熟とリソソームとの融合を阻害する可能性があります14,15。
PD患者では、o-α-synやS129がリン酸化された、α-syn (pS129α-syn) などの修飾されたα-synが有意に増加します。中でも、PDとコントロールを区別する試験では、o-α-syn/t-α-syn比が最も感度が高く (89.3%) 、特異的 (90.6%)です16,17,18,19,20。
Michael J. Fox Foundationは、α-synの凝集体に対して、親和性と特異性が高いPETトレーサーの開発を目指す共同研究施設を設立しました。アミロイドPET検査のように、α-synも可視化できるかどうかは、まだ判明していません。
リソソームの機能不全の研究に用いられる手法
リソソームは、オートファジーやマイトファジーにおいて重要な役割を果たすため、PD患者におけるその形態やエキソサイトーシス、酸性化、位置、運動性、機能などを理解することが、疾患の発症機序のさらなる解明に重要です。抗体技術を用いたリソソームの生物学の研究には様々な方法があり、個々に利点と欠点があります。
ここではその一部の個々の概要を、より良く理解するために有用な科学論文と共に紹介します。
-
Live cell LysoTracker:本手法は、生細胞中のリソソームの数や大きさ、分布を解析することが可能であり、特に治療を目的とする化合物のスクリーニングに有用です。しかし、この方法は、リソソームのpHを測定することはできません。プローブが示す蛍光は、pHにはほとんど依存せず、酸性のオルガネラに無差別に蓄積します。LysoTrackerプローブは、リソソームだけでなく後期エンドソームも標識するため、蛍光シグナルをリソソームpHの指標として用いることはできません。
-
リソソーム拡張プロトコール:メチル基で修飾されたアミノ酸アナログ (LEU-ME) は、リソソームに局在し、リソソーム区画の急速な拡張を誘導する腔内の浸透圧効果を引き起こします。固定後、この技術はリソソームの形態を保持するため、リソソーム寸法の正確な定量と分析に用いることができます。しかし、このプロトコールは短時間の実験にしか適用できません。
-
固定細胞LAMP染色プロトコール:レーザー顕微鏡のある研究室であれば、どの研究室でも実施可能な蛍光顕微鏡によるリソソームの可視化は、リソソームの分布や大きさを調べてその病的な変化の可能性を推測するための、迅速かつ信頼性の高い方法です。LAMP1は、後期エンドソームとリソソームの両方に存在するため、リソソームを特異的に識別するためには、2つ目の抗体マーカーが必要かもしれません。
-
HAタグ標識済みTMEM192を用いたLyso-IP: このリソソーム精製方法は、非常に迅速 (純粋な無傷のリソソームの単離に10分) であり、液体クロマトグラフィーと質量分析 (LC/MS)に適合するバッファーを用います。しかし、リソソームの全体的な回収量は低く、HA-TMEM192を安定して発現する細胞でしか使用できません。
-
Proximity Ligation Assay (PLA法): この技術は、オートファゴソームの開始や伸長、成熟に関与する40種類以上のATGタンパク質の特性評価に用いることができます。この技術は、非常に汎用性が高く、近接した2つのあらゆるタンパク質における多くの解析を定量的に行うことができます。また、パラフィン包埋した組織サンプルや、in vivoの生体組織におけるタンパク質相互作用の研究にも適応可能です。ただし、結果はプローブに用いる抗体の品質に大きく依存し、バッチ特異的抗体の性能により結果にばらつきが生じる可能性があります。オリゴヌクレオチドの非特異的なライゲーションによるバックグラウンドシグナルが生じる可能性もあります。このようなばらつきは、感度と特異性の高い抗体を用いることにより、最小限に抑えることができます。
上記の方法や論文に加えて、本リソースは、リソソームの形態や配置、運動性、機能を解析する方法や、リソソームの研究に現在用いられる方法を幅広く紹介しています。
CSTが提供するパーキンソン病研究用の抗体キット
CSTは、PDにおけるマイトファジーとエンドリソソーム経路の特性評価に有用な、厳選されたAntibody Samplerキットを提供しています。CSTの抗体はすべて、目的のアッセイにおける抗体の機能、特異性、感度を決定するための6つの相補的な戦略からなる、弊社のHallmarks of Antibody Validation (抗体の検証における戦略) を用いて検証されています。
PD研究の前進に役立つCSTのキットをご覧ください:
- Mitophagy Antibody Sampler Kit #43110
- Autophagy Antibody Sampler Kit #4445
- Parkinson's Research Antibody Sampler Kit #8648
参考文献
- Maday S, Holzbaur EL. Autophagosome biogenesis in primary neurons follows an ordered and spatially regulated pathway. Dev Cell. 2014;30(1):71-85. doi:10.1016/j.devcel.2014.06.001
- Lee JJ, Wedow R, Okbay A, et al. Gene discovery and polygenic prediction from a genome-wide association study of educational attainment in 1.1 million individuals. Nat Genet. 2018;50(8):1112-1121. Published 2018 Jul 23 doi:10.1038/s41588-018-0147-3
- McWilliams TG, Prescott AR, Montava-Garriga L, et al. Basal Mitophagy Occurs Independently of PINK1 in Mouse Tissues of High Metabolic Demand. Cell Metab. 2018;27(2):439-449.e5. doi:10.1016/j.cmet.2017.12.008
- Panicker N, Ge P, Dawson VL, Dawson TM. The cell biology of Parkinson's disease. J Cell Biol. 2021;220(4):e202012095. doi:10.1083/jcb.202012095
- Sulzer D, Edwards RH. The physiological role of α-synuclein and its relationship to Parkinson's Disease. J Neurochem. 2019;150(5):475-486. doi:10.1111/jnc.14810
- Fayyad M, Salim S, Majbour N, et al. Parkinson's disease biomarkers based on α-synuclein. J Neurochem. 2019;150(5):626-636. doi:10.1111/jnc.14809
- Kordower JH, Chu Y, Hauser RA, Freeman TB, Olanow CW. Lewy body-like pathology in long-term embryonic nigral transplants in Parkinson's disease. Nat Med. 2008;14(5):504-506. doi:10.1038/nm1747
- Masuda-Suzukake M, Nonaka T, Hosokawa M, et al. Prion-like spreading of pathological α-synuclein in brain. Brain. 2013;136(Pt 4):1128-1138. doi:10.1093/brain/awt037
- Recasens A, Dehay B, Bové J, et al. Lewy body extracts from Parkinson disease brains trigger α-synuclein pathology and neurodegeneration in mice and monkeys. Ann Neurol. 2014;75(3):351-362. doi:10.1002/ana.24066
- Wong YC, Holzbaur EL. Temporal dynamics of PARK2/parkin and OPTN/optineurin recruitment during the mitophagy of damaged mitochondria. Autophagy. 2015;11(2):422-424. doi:10.1080/15548627.2015.1009792
- Winslow AR, Chen CW, Corrochano S, et al. α-Synuclein impairs macroautophagy: implications for Parkinson's disease. J Cell Biol. 2010;190(6):1023-1037. doi:10.1083/jcb.201003122
- Martinez-Vicente M, Talloczy Z, Kaushik S, et al. Dopamine-modified alpha-synuclein blocks chaperone-mediated autophagy. J Clin Invest. 2008;118(2):777-788. doi:10.1172/JCI32806
- Cuervo AM, Stefanis L, Fredenburg R, Lansbury PT, Sulzer D. Impaired degradation of mutant alpha-synuclein by chaperone-mediated autophagy. Science. 2004;305(5688):1292-1295. doi:10.1126/science.1101738
- Tanik SA, Schultheiss CE, Volpicelli-Daley LA, Brunden KR, Lee VM. Lewy body-like α-synuclein aggregates resist degradation and impair macroautophagy. J Biol Chem. 2013;288(21):15194-15210. doi:10.1074/jbc.M113.457408
- Volpicelli-Daley LA, Luk KC, Lee VM. Addition of exogenous α-synuclein preformed fibrils to primary neuronal cultures to seed recruitment of endogenous α-synuclein to Lewy body and Lewy neurite-like aggregates. Nat Protoc. 2014;9(9):2135-2146. doi:10.1038/nprot.2014.143
- Parnetti L, Chiasserini D, Persichetti E, et al. Cerebrospinal fluid lysosomal enzymes and alpha-synuclein in Parkinson's disease. Mov Disord. 2014;29(8):1019-1027. doi:10.1002/mds.25772
- Parnetti L, Farotti L, Eusebi P, et al. Differential role of CSF alpha-synuclein species, tau, and Aβ42 in Parkinson's Disease. Front Aging Neurosci. 2014;6:53. Published 2014 Mar 31 doi:10.3389/fnagi.2014.00053
- Tokuda T, Salem SA, Allsop D, et al. Decreased alpha-synuclein in cerebrospinal fluid of aged individuals and subjects with Parkinson's disease. Biochem Biophys Res Commun. 2006;349(1):162-166. doi:10.1016/j.bbrc.2006.08.024
- Hansson O, Hall S, Ohrfelt A, et al. Levels of cerebrospinal fluid α-synuclein oligomers are increased in Parkinson's disease with dementia and dementia with Lewy bodies compared to Alzheimer's disease. Alzheimer's Res Ther. 2014;6(3):25. Published 2014 May 7. doi:10.1186/alzrt255
- Aasly JO, Johansen KK, Brønstad G, et al. Elevated levels of cerebrospinal fluid α-synuclein oligomers in healthy asymptomatic LRRK2 mutation carriers. Front Aging Neurosci. 2014;6:248. Published 2014 Sep 25 doi:10.3389/fnagi.2014.00248