ストレス顆粒 (SG) は、ストレス反応において形成される重要な細胞の防御機構で、細胞がタンパク質の産生を一時停止し、主要な転写産物を保護するのに役立ちます。通常この顆粒は、ストレスがなくなれば消失します。しかし、このプロセスが破壊されると、タンパク質が集まって有害な凝集体を形成します。

本ブログ記事では、この顆粒がどのように形成され、どのタンパク質がこの顆粒に局在しいて主要なマーカーとして利用できるかについてや、様々な神経変性疾患におけるその役割を紹介します。
ストレス顆粒とは何か?
トマトの細胞で初めて発見されたストレス顆粒は、様々な細胞ストレスに応答して細胞質内に形成される、不規則な形の微小な (0.1 - 4 µm) 構造体です。この顆粒は、タンパク質やリボヌクレオチドの複雑な混合物で構成されており、その形成のプロセスは、酵母からヒトに至るまで多くの生物に共通して保存されています。SGの凝集は、タンパク質の翻訳の中断と翻訳されていないmRNA転写産物の隔離を行い、ストレス応答性の転写産物の翻訳を優先するための、ストレスに対する保護的な適応応答であると考えられています。SGの形成は通常は一過性であり、ストレス要因が取り除かれるとSGは速やかに分解されます。
次のタイムラプスビデオでは、この動的なプロセスの詳細をご覧いただけます1。
NaAsO2ストレス中のストレス顆粒の形成GFP-G3BP1を発現するU-2 OS細胞をNaAsO2 (0.5 mM) に曝露し、20分間撮影しました。画像は、スピニングディスク共焦点顕微鏡上で20秒の間隔で、100X objectiveを使用して取得しました。
SGは生体分子凝縮体のうちの1つであり、非膜オルガネラ (MLO) でもあります。よく研究されている他のMLOとして、核小体やカハール体、核スペックル、プロセシングボディ (P-body) などがあります。MLOは、二重膜を持たない細胞内の分画された領域であり、通常はRNA結合タンパク質 (RBP) とリボヌクレオチドで構成されています。MLOにリクルートされるRBPの多くは、グリシンや非極性残基を多く含む、複雑性の低い天然変性領域 (IDR) を有しています。

IDRは、特有の化学的性質により、他のIDRと数多くの弱い静電的相互作用を持つことができるため、IDRを有するタンパク質が細胞質内で個々の液滴に凝集・分離することを可能にします。この液滴への分離は、一般に液液相分離 (LLPS) と呼ばれ、SGや他の多くのMLOの形成の原動力であると考えられています。LLPSをより分かりやすく理解するために、油がコップ一杯の水の中で液滴に分離する様子を思い浮かべてください。
ストレス顆粒はどのように形成されるのか?
前述したとおり、SGは様々な生物学的および非生物的ストレスに応答して形成され、その結果、eIF2タンパク質の51位のセリンにリン酸化が生じます。この確立されたリン酸化イベントは、ストレスの種類に応じて4つの異なる上流キナーゼ、
-
ウイルス感染のPKR
-
ERストレスのPERK
-
飢餓/栄養の枯渇のGCN2
-
酸化または浸透圧ストレスのHRI2
eIF2は、一度リン酸化するとGDPをGTPに変換できなくなり、タンパク質の翻訳が停止し、40Sリボソーム小サブユニットの表面や翻訳されていないmRNA、翻訳開始因子が露出します。この露出した転写開始前複合体 (PIC) は、SG関連タンパク質をリクルートするための「シード」となり、最終的にSGの凝集を引き起こします3。
腫瘍マーカー:どのようなタンパク質がストレス顆粒に局在するのか?
ストレス顆粒に局在するタンパク質として400種類以上が同定されています4。これらのタンパク質の大半は、ストレスの種類や疾患状態、調査対象の細胞株や組織といった様々な要因により誘導されます。
SGの凝集に必要な主なタンパク質をいくつか紹介します。
G3BP1およびG3BP2
分子量が55 kDaのRBPであるG3BP1は、mRNAの翻訳が停止している部位に最初にリクルートされるタンパク質の1つであり、当初はRasGAPに結合すると考えられていました。しかし現在では、G3BP1自体はSGの凝集の主要なメディエーターではないとしても、重要なメディエーターの1つであることが分かっています。細胞質では、G3BP1はアミノ酸配列の340-415位にあるRNA認識モチーフ (RRM) を介して、翻訳されていないmRNAと相互作用します。
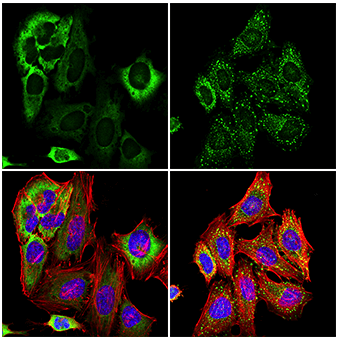 未処理 (左)、あるいは亜ヒ酸ナトリウム処理 (500 μM、30分間、右) したHeLa細胞を、 G3BP1 (E9G1M) XP® Rabbit mAb (緑) を用いて免疫蛍光染色し、共焦点顕微鏡で解析しました。アクチンフィラメントは、 DyLight 554 Phalloidin #13054 (赤) で標識しました。サンプルは、 ProLong Gold Antifade Reagent with DAPI #8961 (青) を用いて封入しました。注釈:血清飢餓処理と亜ヒ酸ナトリウム処理後に、G3BP1の移行とストレス顆粒の形成が生じています。
未処理 (左)、あるいは亜ヒ酸ナトリウム処理 (500 μM、30分間、右) したHeLa細胞を、 G3BP1 (E9G1M) XP® Rabbit mAb (緑) を用いて免疫蛍光染色し、共焦点顕微鏡で解析しました。アクチンフィラメントは、 DyLight 554 Phalloidin #13054 (赤) で標識しました。サンプルは、 ProLong Gold Antifade Reagent with DAPI #8961 (青) を用いて封入しました。注釈:血清飢餓処理と亜ヒ酸ナトリウム処理後に、G3BP1の移行とストレス顆粒の形成が生じています。
G3BP1は、RNAに結合すると、その複数のIDRを介して他の様々なSG関連タンパク質をリクルートして相互作用し、相分離を促進します。一過性のG3BP1の過剰発現は、細胞ストレスがない場合にもSG形成を誘導することから、このプロセスにおいてG3BP1が重要な機能を持つことが分かります5,6。G3BP1の低レベルにより、ストレスへG3BP1の発現量が低下すると、ストレスに応答して形成されるSGの数が減少し、サイズが小さくなります。一方、G3BP1とそのパラログであるG3BP2のダブルノックアウト変異は、ヒ素により誘導されるSG形成を完全に消失させます5,7,8,9。現在、G3BP2については不明なことが多く、SG形成に関してはG3BP1とほぼ重複した機能を持つと考えられています8。
TIA-1およびTIAR
TIA-1とTIARは、通常はSGと関連している、もう一組のRBPです。ストレス誘導により、TIA1/Rは核から細胞質のSGに移行し、そこでG3BP1/2と共にSGの「コア」を形成すると考えられています。TIA1/Rは、翻訳されていないmRNAと相互作用する3つのN末端RRMと、相分離とSG形成に必要なC末端プリオン様ドメイン(PLD)を有しています。11 G3BP1と同様に、TIA1を過剰発現しただけで、ストレスがない状態でのSG形成が誘導されます。10
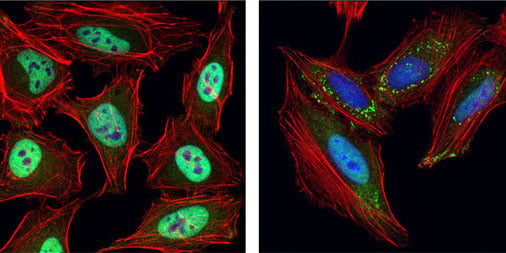
未処理 (左)、あるいはUV処理 (右) したHeLa細胞を、 TIAR (D26E4) Rabbit mAb #8611 (緑) を用いて免疫蛍光染色し、共焦点顕微鏡で解析しました。アクチンフィラメントはDyLight 554 Phalloidin #13054 (赤) で標識しました。DRAQ5 #4084 (DNA蛍光染色試薬) の染色像を青で示しています。UV処理後に、TIARの移行とストレス顆粒の形成が生じています。
その他のタンパク質マーカー
その他にも、FMR1/FMRP、PABPC1、ataxin-2、caprin1、UBAP2Lなどの多くのタンパク質がSGにリクルートされ、SGの調節に関与していることが分かっています。UBAP2Lは、G3BP1/2の上流でSG形成に関与している可能性があるため、特に注目されています。この新しい発見は、SG形成の動的さについてさらなる研究が必要であることを示しています13。
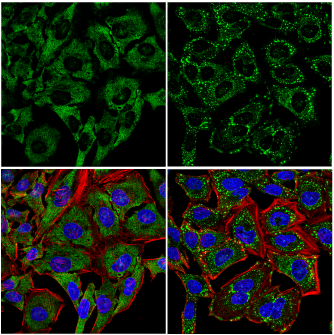
未処理 (左)、あるいは亜ヒ酸ナトリウム処理 (500 μM、30分間、右) したHeLa細胞を、UBAP2L (E5X4E) Rabbit mAb (緑) を用いて免疫蛍光染色し、共焦点顕微鏡で解析しました。アクチンフィラメントはDyLight 554 Phalloidin #13054 (赤) で標識しました。DRAQ5 #4084 (DNA蛍光染色試薬) の染色像を青で示しています。亜ヒ酸ナトリウム処理後に、UBAP2Lの移行とストレス顆粒の形成が生じています。
神経変性疾患におけるストレス顆粒
正常な状態では、ストレス顆粒の形成は一過性の可逆的なプロセスであり、ストレスが取り除かれると、VCPやSQSTM1などの複数のオートファジー促進タンパク質のリクルートによって、SGの分解が行われます。しかし、SGはタンパク質間の相互作用を非常に多く含むため、分解が阻害されると、有害なタンパク質凝集体を形成する恐れがあります。このように、SGが可溶性の液相混合物から不溶性の固相凝集体へと移行することにより、様々な神経変性疾患の特徴である細胞質封入体の形成のシードとして機能する可能性があります。
転写の調節とエクソンのスプライシングに関与するRNA結合タンパク質であるTDP43は、筋萎縮性側索硬化症 (ALS) に関連する主要な病原性タンパク質です。また、TDP43の蓄積は、アルツハイマー病 (AD) や前頭側頭型認知症 (FTD) とも関連しています。TDP43の疾患関連変異は、TDP43の核から細胞質のストレス顆粒への誤った局在化やSGのサイズと数の増加、SGの病原性封入体への変換など、SGの生物物理的特性における無数の変化を促進します14,15,16,17。
FUSは、転写調節やmRNA前のスプライシング、DNAの損傷への応答の役割を担う、もう1つのRNA結合タンパク質です。TDP43と同様に、FUSを含むタンパク質凝集体の病的な蓄積は、いくつかの神経変性疾患において確認されています。FUSの変異は、ALSとFTDの患者に多く観察されており、通常はタンパク質のC末端にある核移行シグナル配列に生じます。これらの変異は、主に核内に存在するタンパク質の細胞質への誤った局在化を引き起こし、その結果、細胞質ストレス顆粒へのFUS凝集体の蓄積を促進します18。
従来のRBPとは異なりますが、微小管関連タンパク質TauもRNAに結合し、TIA1、Ataxin-2、PABPC1などの複数のSG関連タンパク質と共局在することが示されています。9,20。TauにもLLPSが生じ、RNAの存在下ではより増強されますが、TauはSGから独立してRNA顆粒を形成することはありません。これは、TauがmRNAやRBPとの相互作用を介してSGにリクルートされ、そこで不溶性のTau凝集体を形成する可能性があることを示唆しています。Tauの過剰なリン酸化も、SGへのリクルートに影響を与える可能性があります20。
今後の展望
ストレス顆粒は、一連のシグナル伝達イベントと液液相分離を介して、細胞がタンパク質の翻訳の変化を起こすための、独特で大変興味深い生理的な適応応答です。熱心に研究されている分野であり、今後の研究により、SGの形成・分解、これらの構造体にリクルートされる新たな構成因子、ヒト疾患との関連性などが解明されることが期待されます。ALSやFTDなどの神経変性疾患とSGの関連性の研究は大きく進展しており、SGタンパク質の誤った折り畳みや凝集を標的とする治療法が、これらの疾患の治療に役立つ可能性があります。
追加リソース:
- ストレス顆粒のさらなる情報は、ストレス顆粒のライフサイクルのインタラクティブ概略図をご覧ください
- その他のMLOについては、生体分子凝縮体マーカーのポスターをダウンロードしてご覧ください
- アルツハイマー病のインタラクティブパスウェイ図をご覧ください
- パンフレット:神経変性の特性:基礎となる生物学的プロセスの特定をダウンロードしてご活用ください
- Scienceウェビナー、ストレス顆粒:神経変性との関連性を解読するをご覧ください
参考文献:
- Wheeler JR, Matheny T, Jain S, Abrisch R, Parker R. Distinct stages in stress granule assembly and disassembly. Elife. 2016;5:e18413. Published 2016 Sep 7. doi:10.7554/eLife.18413. CC BY 4.0.
- Mahboubi H, Stochaj U. Cytoplasmic stress granules: Dynamic modulators of cell signaling and disease. Biochim Biophys Acta Mol Basis Dis. 2017;1863(4):884-895. doi:10.1016/j.bbadis.2016.12.022
- Panas MD, Ivanov P, Anderson P. Mechanistic insights into mammalian stress granule dynamics. J Cell Biol. 2016;215(3):313-323. doi:10.1083/jcb.201609081
- Marcelo A, Koppenol R, de Almeida LP, Matos CA, Nóbrega C. Stress granules, RNA-binding proteins and polyglutamine diseases: too much aggregation?. Cell Death Dis. 2021;12(6):592. Published 2021 Jun 8. doi:10.1038/s41419-021-03873-8
- Tourrière H, Chebli K, Zekri L, et al. The RasGAP-associated endoribonuclease G3BP assembles stress granules. J Cell Biol. 2003;160(6):823-831. doi:10.1083/jcb.200212128
- Reineke LC, Dougherty JD, Pierre P, Lloyd RE. Large G3BP-induced granules trigger eIF2α phosphorylation. Mol Biol Cell. 2012;23(18):3499-3510. doi:10.1091/mbc.E12-05-0385
- Ghisolfi L, Dutt S, McConkey ME, Ebert BL, Anderson P. Stress granules contribute to α-globin homeostasis in differentiating erythroid cells. Biochem Biophys Res Commun. 2012;420(4):768-774. doi:10.1016/j.bbrc.2012.03.070
- Matsuki H, Takahashi M, Higuchi M, Makokha GN, Oie M, Fujii M. Both G3BP1 and G3BP2 contribute to stress granule formation. Genes Cells. 2013;18(2):135-146. doi:10.1111/gtc.12023
- Kedersha N, Panas MD, Achorn CA, et al. G3BP-Caprin1-USP10 complexes mediate stress granule condensation and associate with 40S subunits [published correction appears in J Cell Biol. 2020 Jan 6;219(1):]. J Cell Biol. 2016;212(7):845-860. doi:10.1083/jcb.201508028
- Kedersha NL, Gupta M, Li W, Miller I, Anderson P. RNA-binding proteins TIA-1 and TIAR link the phosphorylation of eIF-2 alpha to the assembly of mammalian stress granules. J Cell Biol. 1999;147(7):1431-1442. doi:10.1083/jcb.147.7.1431
- Waris S, Wilce MC, Wilce JA. RNA recognition and stress granule formation by TIA proteins. Int J Mol Sci. 2014;15(12):23377-23388. Published 2014 Dec 16 doi:10.3390/ijms151223377
- Gilks N, Kedersha N, Ayodele M, et al. Stress granule assembly is mediated by prion-like aggregation of TIA-1. Mol Biol Cell. 2004;15(12):5383-5398. doi:10.1091/mbc.e04-08-0715
- Cirillo L, Cieren A, Barbieri S, et al. UBAP2L Forms Distinct Cores that Act in Nucleating Stress Granules Upstream of G3BP1. Curr Biol. 2020;30(4):698-707.e6. doi:10.1016/j.cub.2019.12.020
- Neumann M, Sampathu DM, Kwong LK, et al. Ubiquitinated TDP-43 in frontotemporal lobar degeneration and amyotrophic lateral sclerosis. Science. 2006;314(5796):130-133. doi:10.1126/science.1134108
- Winton MJ, Igaz LM, Wong MM, Kwong LK, Trojanowski JQ, Lee VM. Disturbance of nuclear and cytoplasmic TAR DNA-binding protein (TDP-43) induces disease-like redistribution, sequestration, and aggregate formation. J Biol Chem. 2008;283(19):13302-13309. doi:10.1074/jbc.M800342200
- Bentmann E, Neumann M, Tahirovic S, Rodde R, Dormann D, Haass C. Requirements for stress granule recruitment of fused in sarcoma (FUS) and TAR DNA-binding protein of 43 kDa (TDP-43). J Biol Chem. 2012;287(27):23079-23094. doi:10.1074/jbc.M111.328757
- Mann JR, Gleixner AM, Mauna JC, et al. RNA Binding Antagonizes Neurotoxic Phase Transitions of TDP-43. Neuron. 2019;102(2):321-338.e8. doi:10.1016/j.neuron.2019.01.048
- Vance C, Scotter EL, Nishimura AL, et al. ALS mutant FUS disrupts nuclear localization and sequesters wild-type FUS within cytoplasmic stress granules. Hum Mol Genet. 2013;22(13):2676-2688. doi:10.1093/hmg/ddt117
- Vanderweyde T, Apicco DJ, Youmans-Kidder K, et al. Interaction of tau with the RNA-Binding Protein TIA1 Regulates tau Pathophysiology and Toxicity. Cell Rep. 2016;15(7):1455-1466. doi:10.1016/j.celrep.2016.04.045
- Wolozin B, Ivanov P. Stress granules and neurodegeneration. Nat Rev Neurosci. 2019;20(11):649-666. doi:10.1038/s41583-019-0222-5